 Получите свидетельство
Получите свидетельство Вход
Вход

Оглавление.
1. Гетеротрофный способ питания растений…………….............................2 стр.
1.1.Общая характеристика гетеротрофных растений.
1.2.Сапрофиты.
1.3.Паразиты.
1.4. Насекомоядные растения.
1.5. Пищеварение у растений.
2. Физиология прорастания семян.
2.1. Фазы прорастания семян.
2.2. Морфология прорастающего семени.
Приложение 1.
Приложение 2.
Список литературы.
1. Гетеротрофный способ питания растений
1.1.Общая характеристика гетеротрофных растений.
К гетеротрофам относят всех животных, грибы и большинство бактерий. Среди растений также имеются факультативные или облигатные гетеротрофы, получающие органическую пищу из внешней среды – сапрофиты, паразиты и насекомоядные растения. Сапрофиты (сапротрофы) питаются органическими веществами разлагающихся остатков растений и животных, паразиты – органическими веществами живых организмов. Насекомоядные растения способны улавливать и переваривать мелких беспозвоночных. Однако в жизни всех растений есть периоды, когда они питаются только за счет запасенных ранее органических веществ. К таким периодам относят прорастание семян, органов вегетативного размножения (клубней, луковиц и др), рост побегов из корневищ, развитие почек и цветков. Все ткани и органы растения гетеротрофно питаются в темноте (выращивание изолированных тканей без света на органоминеральной среде). Таким образом, гетеротрофное питание клеток и тканей становится обычным, как и фотосинтез.
Гетеротрофный способ питания – усвоение как низкомолекулярных органических соединений, так и высокомолекулярных (белков, жиров, углеводов), но они должны пройти обработку – пищеварение.
1.2. Сапрофиты.
Механизмы сапрофитного питания растений и грибов сходны. В плазмолемме гиф грибов функционирует Н+-помпа (водородная помпа), с помощью которой в окружающую среду выделяются кислые гидролазы. Это приводит к гидролизу сложных органических соединений, которые затем всасываются грибом. Механизм всасывания также связан с работой Н+-помпы в плазмолемме. При закислении наружной примембранной зоны снижается диссоциация органических кислот и они проникают в клетки в виде нейтральных молекул. Этот способ распространен у водорослей (диатомовые водоросли, живущие на глубине, куда не проникает свет, питаются органическими веществами из окружающей среды). При большом количестве растворимых органических веществ в водоемах к гетеротрофному питанию переходят хлорококковые, эвгленовые и др водоросли.
У покрытосеменных растений сапрофитный способ питания встречается редко. Эти растения не имеют или имеют мало хлорофилла и не способны к фотосинтезу. Для построения своего тела они используют гниющие остатки растений и животных. Gidiophytum formicarum – полукустарник, стебель которого образует крупный клубень, пронизанный многочисленными ходами, в которых поселяются муравьи. {Приложение 1рис.1} Растение использует в пищу продукты жизнедеятельности муравьев. Меченые личинки мух были переварены растением через месяц.
Некоторые виды образуют симбиоз с грибами – это микотрофные растения. На ранних этапах все орхидные вступают в симбиоз с грибами, так как запасных питательных веществ в их семенах недостаточно для роста зародыша. Гифы грибов проникают в семена и поставляют зародышу органические и минеральные вещества. У взрослых орхидей гифы грибов внедряются в периферическую зону корней, но дальше проникнуть не могут. Их дальнейшему росту препятствует фунгистатическое действие клеток глубинных тканей корня, а также слой довольно больших клеток с крупными ядрами, похожих на фагоциты. Эти клетки способны переваривать гифы грибов и усваивать освобождающиеся органические вещества. У подъельника (Monotropa), тоже симбиоз с грибом. Причем во многих случаях эти симбиотические отношения можно рассматривать как форму паразитизма, Когда клетки растений переваривают гифы гриба, проникшие в клетки корня. Таким образом, сапрофитом является гриб, а высшее растение паразитирует на нем. Гифы гриба могут соединять корень подъельника с корнями дерева, и тогда подъельник становится паразитом, получающим органические в. Паразиты поглощают из тела хозяина в основном сахарозу, глутаминовую и аспарагиновую кислоты и их амиды, вещества от другого растения.
1.3. Паразиты
Микоризный гриб может выступать как паразит. Гифы гриба образуют выросты-гаустории. плотно прилегающие или проникающие в корень. Они высасывают питательные вещества (прежде всего углеводы) из растения.
Высшие растения-паразиты - это высокоспециализированные однолетники или многолетники с редуцированными или полностью утраченными в ходе эволюции листьями и корнями. Имеются виды, совершенно лишенные хлорофилла и не способные к фотосинтезу. Заразиха паразитирующая на корнях культурных растений.{Приложение 1 Рис. 2} Ее семена прорастают лишь под влиянием корневых выделений растения-хозяина. Как только кончик зародышевого корня проростка соприкоснется с корнем хозяина, он преобразуется в гаусторию (присоску), начинающую выделять гидролазы, растворяющие клеточные стенки, и активно внедряющуюся в корень. Заразиха поглощает большое количество азотистых веществ, углеводов и минеральных элементов, особенно фосфора, а также воды из корней растений-хозяев.
У вьющегося паразитного травянистого растения повилики нитевидные стебли с редуцированными листьями-чешуйками обвиваются вокруг стеблей растений-хозяев и присасываются к ним с помощью гаусторий. Гаустории повилики - преобразованные придаточные корни, принимающие форму диска. Клетки из центральной части диска внедряется в коровую паренхиму растения-хозяина и достигает центрального цилиндра, откуда повилика получает воду, органические вещества и минеральные элементы. Проростки повилики, совершая ростовые вращательные движения, находят растение-хозяина, реагируя на градиент влажности и выделяемые им вещества.
Раффлезия питается соками корней тропических лиан. В тело хозяина она внедряется с помощью гаусторий, выделяющих ферменты, разрушающие клеточные стенки. Всю свою жизнь раффлезия проводит в теле хозяина - под землей. Лишь ее цветки (диаметр 1,5 м, красного цвета с запахом гниющего мяса) появляются на поверхности почвы.
Паразиты поглощают из тела хозяина в основном сахарозу, глутаминовую и аспарагиновую кислоты и их амиды.
1.4. Насекомоядные растения.
В настоящее время известно свыше 400 видов покрытосеменных насекомоядных растений. Они ловят мелких насекомых и другие организмы, переваривают и используют как дополнительный источник питания. Большинство из них встречается на бедных азотом болотистых почвах, есть эпифитные и водные формы. Листья насекомоядных растений трансформированы в специальные ловушки, выполняющие и функцию фотосинтеза. По способу ее ловли растения делят на две группы. 1) Пассивная ловля, добыча а) прилипает к листьям, желёзки которых выделяют липкую слизь, содержащую кислые полисахариды (библис, росолист), или б) попадает в специальные ловушки в виде кувшинов, урн, трубочек, окрашенных в яркие цвета и выделяющие сладкий ароматный секрет (саррацения, дарлингтония).
Микориза большинством растений используется главным образом для увеличения поглощения воды и минеральных солей.
2) Активный захват насекомых а) приклеивание добычи липкой слизью и обволакивание ее листом или волосками (жирянка, росянка), б) ловля по принципу капкана - с захлопыванием ловчих листьев над добычей (альдрованда, венерина мухоловка), в) ловчие пузырьки, в которые насекомые втягиваются с водой благодаря поддерживаемому в них вакууму (пузырчатка).
Общим для всех ловчих приспособлений является привлечение насекомых с помощью полисахаридных слизей или ароматного секрета (нектара), выделяемых или самими ловчими аппаратами, или желёзками вблизи от ловушки. Быстрые движения ловчих органов осуществляются путем изменений в них тургора в ответ на раздражение чувствительных волосков, вызванное движениями насекомого.
1.5. Пищеварение у растений.
Различают три типа пищеварения: внутриклеточное, мембранное и внеклеточное.
Внутриклеточное — самый древний тип пищеварения. У растений оно происходит не только в цитоплазме, но и в вакуолях, пластидах, белковых телах, сферосомах.
Мембранное пищеварение осуществляется ферментами, локализованными в клеточных мембранах, что обеспечивает максимальное сопряжение пищеварительных и транспортных процессов. Оно хорошо изучено в кишечнике ряда животных. У растений мембранное пищеварение не исследовалось.
Внеклеточное пищеварение происходит тогда, когда гидролитические ферменты, образующиеся в специальных клетках, выделяются в наружную среду и действуют вне клеток. Этот тип пищеварения характерен для насекомоядных растений; он осуществляется и в других случаях, в частности в эндосперме зерновок злаков.
2.Физиология прорастания семян.
Прораста́ние семя́н, или гермина́ция (лат. germinatio) — переход семян растений от покоя к активной жизнедеятельности, начальный этап онтогенеза растений, на котором образуется проросток. Происходит при обеспеченности влагой и кислородом, подходящем температурном и световом режиме. В процессе прорастания повышается обмен веществ в зародыше и эндосперме; семена набухают в воде, крахмал, жиры и белки распадаются на сахар, жирные кислоты и аминокислоты. Обычно первым прорастает корешок, далее — гипокотиль или эпикотиль (у разных растений).
В случае недостатка кислорода накапливаются вредные для зародыша вещества — этиловый спирт, молочная кислота, аммиак; при недостатке температуры снижается поступление воды в семена и активация обмена веществ, нарушается соотношение различных регуляторов роста. Некоторые из семян не прорастают, находясь в подходящих условиях, из-за твёрдости покровов и не выхода из состояния покоя; в этом случае возможно механическое повреждение покровов.
2.2. Фазы прорастания семян. ( Приложение 2 рис. 1)
Период прорастания состоит из последовательных этапов – фаз прорастания. Каждой фазе присуща определенная продолжительность, определенные биохимические и морфологические изменения, происходящие в семени, а также определенные требования к условиям среды.
Фазы прорастания семян:
водопоглощение;
набухание и наклёвывание;
рост первичных корешков;
развитие ростка;
становление проростка.
А.Фаза водопоглощения.
Сухие семена, находящиеся в состоянии покоя, поглощают воду из воздуха (если его относительная влажность больше 75%) или из какого-либо субстрата до наступления критической влажности, которая является строго определенной величиной для каждой культуры. Поступающую воду поглощают гидрофильные коллоиды семени. Вода включается в содержимое клетки, где связывается различными ее соединениями, и поэтому в семени не происходит заметной активизации биохимических процессов и не наблюдается никаких изменений в морфологии. Поглощение воды может несколько повысить интенсивность дыхания семян (к концу фазы в 2–3 раза), но общий ее уровень остается очень низким.Основа фазы водопоглощения – физико-химическое явление, сорбция.
Продолжительность фазы зависит от состояния семян, температуры и влажности субстрата, с которым соприкасается семя. Довольно большая продолжительность, если влага поступает из воздуха, очень короткая, если семена находятся в воде. Однако в последнем случае для равномерного насыщения клеток требуется еще некоторое время, и только после такого распределения влаги наступает вторая фаза.
B. Фаза набухания семян
Начинается с момента появления в семенах свободной влаги. Она активизирует жизнедеятельность клеток, усиливает гидролитические процессы, переводит в активное состояние ферментную систему, ведет к перестройке коллоидов. При этом дыхательный коэффициент увеличивается в сотни и даже в тысячи раз. Заканчивается фаза наклёвыванием.
Молекулы воды проникают в среду высокомолекулярных соединений и раздвигают отдельные звенья в цепи их молекул. Все это вызывает не только ослабление самих цепей молекул, но и сопровождается гидролизом последних, что ведет к интенсификации всех жизненных процессов. В процессе набухания семян оболочки их приобретают эластичность, а само семя увеличивается в объеме.
Процесс набухания семян может характеризоваться двумя показателями: 1) степенью набухания – это количество воды в граммах, поглощенное семенами в фазе набухания в пересчете на 1 г сухого вещества; 2) числом набухания – количество воды в миллилитрах, которое поглощается 1 мл сухого вещества семени. Иногда характеризуют процесс набухания давлением, которое возникает в результате увеличения объема при набухании. Это так называемое давление набухания достигает нескольких сот атмосфер и также является характерным для каждого вида. Прирост веса в результате поглощения воды и прирост объема нарастают неодинаковыми темпами – обычно увеличение объема идет быстрее, и оно скорее заканчивается, чем прирост веса.
Фаза набухания заканчивается поглощением определенного количества воды, которое обеспечивает протекание всех жизненных процессов, связанных с прорастанием. В зависимости от химического состава семян и их природы требуется разное количество воды для наклевывания семян.
Набухание прекращается вследствие полного насыщения клеток или из-за наступления равновесия между поступлением воды в семена и диффузией растворимых веществ из него. Для нормального протекания этой фазы требуется определенная температура, влажность и кислород. При подсыхании наклюнувшихся семян возможно возвращение в предыдущую фазу, исходную.
C.Фаза роста первичных корешков
Начинается с момента деления клеток первичного корешка, но морфологически ее можно зафиксировать несколько позже – при появлении над оболочкой семени первичного корешка. В этой фазе также происходит новая качественная перестройка биохимических процессов, подготавливающая условия для возможности роста ростка (в корнях синтезируются витамины и т.п.). Для нормальной биохимической перестройки и роста корешков требуется иной гидротермический режим, чем для протекания других фаз. Заканчивается фаза готовностью семени к развитию ростка.
Для большинства культур еще возможно прекращение прорастания семян на этой фазе и возвращение их в исходное состояние (состояние покоя), хотя для некоторых культур такой переход связан уже с нарушением физиологии и морфологии прорастания.
D. Фаза развития ростка
Начинается с появлением ростка и заканчивается переходом проростка на автотрофное питание. Продолжается дальнейший рост корешков, но уже имеются все возможности и для развития ростка, который также интенсивно растет. Но здесь уже требуются иные условия питания и внешней среды.Из этой фазы уже нет возврата в состояние покоя, и при подсыхании развивающееся семя гибнет. Фаза оканчивается с появлением у проростка сформированного колеоптиле у злаков или с формированием почечки у других культур.
E. Становление проростка.
На этой фазе и заканчивается процесс прорастания семян, но молодой проросток еще является объектом семеноведческих исследований. Развивающийся проросток находится в сложной зависимости от экологических условий, но еще получает от семени основное питание и некоторые специфические соединения.
Проросшими семенами следует считать только те, которые имеют сформированный проросток (появление ростка с первичными корешками), если нет ростка, то независимо от длины корешков семена нельзя назвать проросшими, а только прорастающими (то есть находящимися на разных фазах прорастания). Для всех остальных культур проросшими семенами считаются такие, которые имеют корешок, равный не менее длины семени, а для круглых семян – не менее диаметра семени.
2.2.Морфология прорастающего семени.
Первым видимым морфологическим признаком прорастания семян является наклевывание, а затем появление корешка. Корешок растет в длину благодаря тому, что у него на конце происходит бурное деление клеток, которые образуют ростовую зону корешка (колеориза), а сам кончик корешка прикрывается корневым чехликом – утолщением разной формы, выполняющим защитные функции.
Как только развивающийся корешок достигает семенной кожуры, он разрывает ее вблизи микропиле и выходит наружу. Если семя заключено, кроме того, в плодовую оболочку, то корешок пробивает и ее. У семян, имеющих эндосперм, корешок обычно очень тонкий, у других семян относительно толще. Как правило, у всех культур появляется один корешок, но у злаковых, кроме главного корешка, очень скоро из придаточных почек развиваются еще боковые или придаточные корешки. Семена полевых культур имеют следующее количество корешков: пшеница озимая от 2 до 6 (корешков больше у крупнозерных сортов и меньше у мелкозерных); пшеница яровая от 3 до 7 (в среднем 5–6); рожь озимая от 4 до 9 (в среднем 5–6); ячмень шестирядный 5–6, двурядный 7–8; овес от 2 до 6 (в среднем 3–4). Кукуруза просо, могар и чумиза прорастают только одним корешком.
Число зародышевых корешков может характеризовать качество семян. Имеются данные, показывающие огромную роль первичных корешков в снабжении растения водой и в формировании урожая, поэтому семеноведам следует обратить большое внимание на изучение зародышевых корешков. Корешки начиная от чехлика и до семени или до гипокотиля (у бобовых) покрыты многочисленными волосками, которые снабжают корень водой и питательными веществами. Признак нормального развития корешка – наличие свежих волосков и проявление геотропизма, то есть изгиба корешка.
У двудольных культур после появления зародышевого корешка растет подсемядольное колено (гипокотиль), выносящее семядоли зародыша вместе с почечкой, расположенной между ними, на поверхность почвы (Приложение 2. рис.2). Из этой почки образуется стебелек и первые листья, по которым часто распознают виды (особенно у крестоцветных).
У гороха, вики, конского боба и некоторых других бобовых растений после появления корешка растет надсемядольное колено (эпикотиль), а семядоли остаются в почве (Приложение 2. рис.2)
Приложение 1.
Рисунок 1. Hydnophytum FORMICARUM - Хиднофитум Формикарум, Хиднофитум муравьиный.

Рисунок 2. Заразиха паразитирующая. (Orobanche)

Рисунок 3. Пузырчатка обыкновенная (Utricularia vulgaris)

Приложение 2
Рисунок 1. Прорастание семени фасоли.
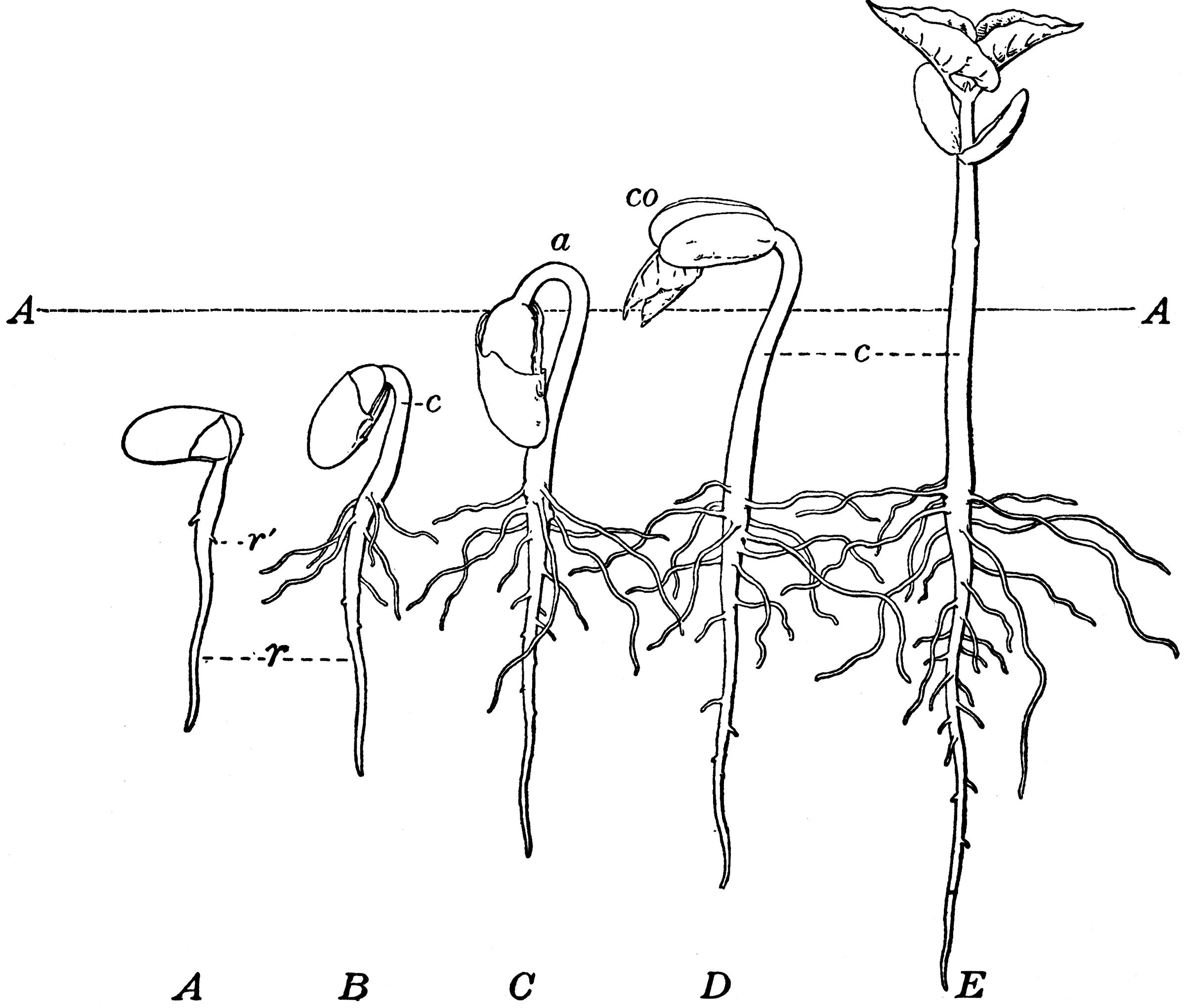
Рисунок 2.Типы прорастания семян. (однодольные и двудольные).

Список литературы.
1.Лекции по курсу «Физиология растений и основы ботаники».
2..https://studizba.com/lectures/biologicheskie-discipliny/kornevoe-i-geterotrofnoe-pitanie/1639-geterotrofnyy-sposob-pitaniya-rasteniy.html
3. https://studfile.net/preview/16531687/page:13/
4.https://rosselhoscenter.ru/ob-uchrezhdenii/filialy/privolzhskiy/kirovskaya-oblast/fiziologiya-prorastaniya-semyan/
5. https://ru.wikipedia.org/wiki/Прорастание_семян









 Методический материал (1.33 MB)
Методический материал (1.33 MB)
 0
0 396
396 0
0 Нравится
0
Нравится
0






