 Получите свидетельство
Получите свидетельство Вход
Вход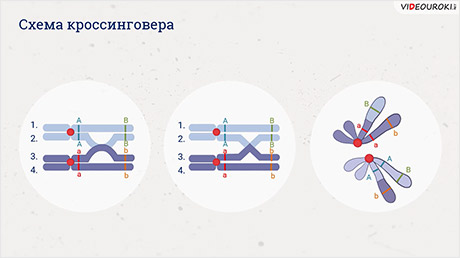



Моногибридное скрещивание заключается в скрещивании организмов, которые отличаться только по одному признаку.

Мендель также работал над скрещиванием организмов, которые отличаются по двум признакам.
Такое скрещивание называют дигибридным. Если скрещивать организмы, которые отличаются по трём признакам, то такое скрещивание называют тригибридным. Скрещивание особей, которые отличаются по нескольким признакам называют полигибридным.
Гибриды, гетерозиготные по двум парам генов, называют дигетерозиготными, а в случае отличия их по трём и многим генам — три- и полигетерозиготными соответственно.
Грегор Мендель приступил к изучению результатов дигибридного скрещивания после того, как установил закономерности моногибридного скрещивания.
Он исследовал характер расщепления при скрещивании двух чистых линий гороха, которые отличались цветом семян (жёлтые и зелёные) и их формой (гладкие и морщинистые). При таком скрещивании признаки определяются различными парами генов: одна пара генов отвечает за цвет семян, другая – за их форму. При этом аллель жёлтой окраски горошин (А-большое), доминирует над зелёной (а-малое). А аллель гладкой формы семян (B-большое) – над морщинистой (b-малое).
По закону единообразия гибридов первого поколения семена гороха (дигибридные семена) в поколении (F1) были жёлтыми и гладкими.
А генотип гибридов первого поколения —А-большое а-малое Бэ-большое – бэ-малое, то есть является дигетерозиготным.
На второй год после самоопыления гибридов первого поколения во втором поколении в соответствии с законом расщепления вновь появились морщинистые и зелёные семена.
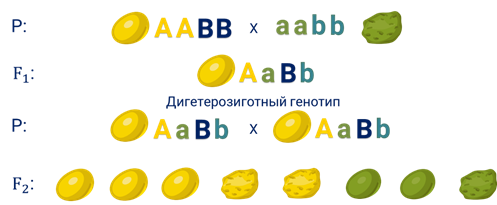
Оказалось, что 315 горошин были жёлтыми гладкими, 101 жёлтыми морщинистыми, 108 зелёными гладкими и 32 зелёными морщинистыми. Соотношение фенотипов очень близко к соотношению 9:3:3:1.
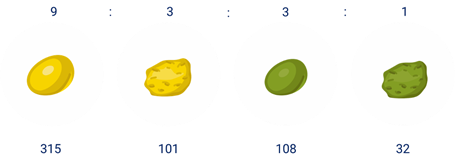
Для того чтобы понять, каким образом комбинируются при скрещивании двух гибридов первого поколения все возможные гаметы, английским генетиком Реджинальдом Пеннетом была предложена решётка, которая впоследствии получила название «решётка Пеннета».
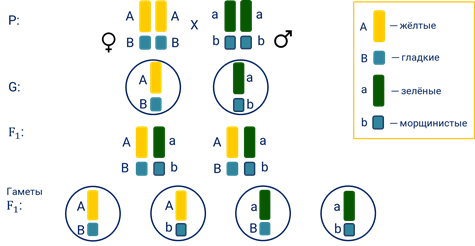
Она позволяет наглядно представить все виды комбинаций генов в гаметах и результаты их слияния.
Вдоль одной стороны решётки располагают мужские гаметы, вдоль другой — женские. А в клетках таблицы на пересечении строк и колонок записывают генотипы потомства в виде комбинаций этих гамет.
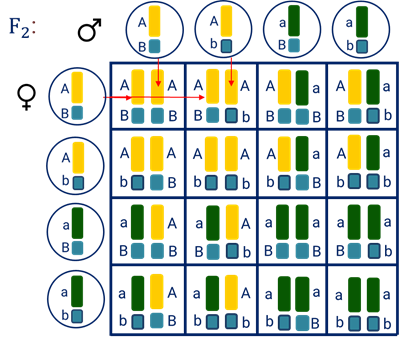
В результате слияния четырёх видов гамет, возникающих у растений из первого поколения (F1) (А-большое Бэ-большое, А-большое бэ-малое, а-малое Бэ-большое, а-малое бэ-малое), во втором поколении (F2) возникает девять различных генотипов.
Но эти девять генотипов проявляются в виде четырёх фенотипов: жёлтые-гладкие, жёлтые-морщинистые, зелёные-гладкие и зелёные морщинистые семена.
Причём соотношение фенотипов составляет 9:3:3:1. Если подсчитать, каким будет соотношение между жёлтыми и зелёными горошинами, то оно будет равняться 3:1. Такое же соотношение будет и между гладкими и морщинистыми семенами.
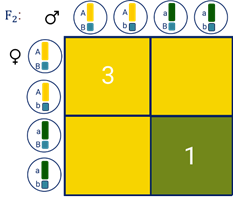
Расщепление при дигибридном скрещивании представляет собой два независимо идущих моногибридных расщепления, которые как бы накладываются друг на друга.
Закон независимого наследования признаков −третий закон Менделя
Мендель обнаружил что расщепление по разным признакам происходит независимо.
В этом сущность третьего закона Менделя — закона независимого наследования признаков, или независимого комбинирования генов.
Он формулируется так: при дигибридном скрещивании гены и признаки, за которые эти гены отвечают, наследуются независимо друг от друга.
Это означает, что при образовании материнских и отцовских гамет в каждую из них может попасть любой аллель из одной пары вместе с любым другим из другой пары.
Следует помнить, что данный закон справедлив в тех случаях, когда гены рассматриваемых признаков располагаются в разных гомологичных хромосомах.

Таким образом Мендель, не имея представления о генах показал механизмы наследования признаков, изучая закономерности, по которым признаки передаются из поколения в поколение.
Рассмотрим закон независимого наследования признаков с позиции цитологии с уже имеющимися на сегодняшний день знаниями.
Выделим гомологичные хромосомы с генами, влияющими на окраску семян. В другой паре гены влияющие на развитие формы горошин. Далее идут 2 мейотических деления мейоз один и мейоз два.
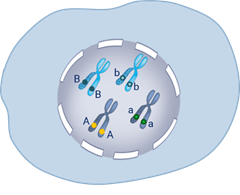
В результате делений образуются 4 типа гамет. В одной оказались гены (А-большое – Бэ-большое) в данном случае это гены жёлтой окраски и гладкой формы семян.
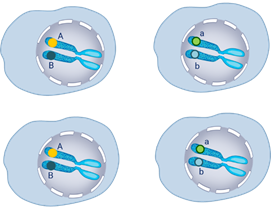
Ещё одна гамета, она отличается от первой геном морщинистой формы (А-большое бэ-малое). То есть содержит рецессивный ген.

Гены могут комбинироваться и в таких сочетаниях (Бэ-большое – а -малое) и (а-малое бэ-малое).
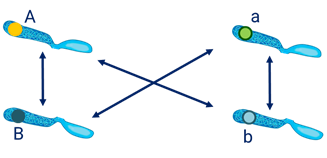
Аналогичные комбинации генов возникают и в мужских половых клетках. Такие сочетания получились потому что гены находились в разных хромосомах.
Что же будет если признаки будут располагаться в одной хромосоме?
После открытых Менделем законов в 1906 году учёные Уильям Бэтсон и Реджинальд Пэннет обнаружили, что в некоторых случаях расщепления по правилам Менделя не происходит.
Например, у душистого горошка два признака – форма пыльцы и окраска цветков – не дают независимого расщепления в потомстве в соотношении 3:1 (три к одному) и потомки остаются похожи на родительской особи. При последующем анализе оказалось, что гены этих двух признаков лежат в одной хромосоме.
Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался американский генетик Томас Морган.
Он предложил закон сцепленного наследования (закон Моргана): гены, которые находятся в одной хромосоме, при мейозе попадают в одну гамету, то есть наследуются сцеплено.
Что значит сцеплено? Это значит, что гены находятся в одной хромосоме. И между ними возникают силы сцепления, то есть силы взаимодействия. И чем ближе эти гены, тем сильнее взаимодействие.
Каковы же принципы наследования генов, расположенных в одной хромосоме?
Вспомним что Мендель проводил свои опыты на горохе. При дигибридном скрещивании во втором поколении соотношение между жёлтыми и зелёными горошинами составляло 3:1.
У Моргана основными объектами для изучения были фруктовые мушки дрозофиллы. В природе они разводятся на опавших и прелых фруктах, из-за чего и получили своё название. Дрозофила имеет диплоидный набор хромосом равный 8.
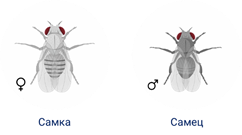

Морган скрещивал двух мушек самка была серой с длинными крыльями, а самец с черным телом и зачаточными крыльями. В первом поколении особь имела серое тело и длинные крылья.

Значит ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обуславливающий развитие нормальных крыльев, — над геном недоразвитых (зачаточных) крыльев.
При дальнейшем скрещивании между собой гибридных мух первого поколения во втором поколении появляются два фенотипа вместо четырёх ожидаемых.
Это позволило сделать вывод, что гены окраски тела и длины крыльев дрозофил находятся в одной хромосоме.
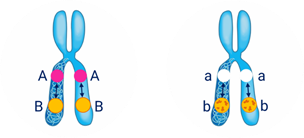
Обратите внимание на запись при сцепленном наследовании.
Символы А-большое Бэ-большое А-большое Бэ-большое и а-малое бэ-малое а-малое бэ-малое располагаются не рядом как мы записывали их ранее, а как бы друг под другом с двумя чёрточками. Чёрточками мы условно обозначаем хромосомы.
Однако в этих же опытах было описано отклонение от закона Моргана. При скрещивании дигетерозиготной особи, которая получилась в первом поколении и чёрной мушки с зачаточными крыльями, имеющая оба рецессивных признака. Получился необычный результат.

Среди гибридов второго поколения было небольшое число особей с перекомбинацией тех признаков, гены которых лежат в одной хромосоме.
В потомстве явно преобладали особи с признаками родительских форм (41,5% были серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% были серые с зачаточными крыльями и 8,5% — черные длиннокрылые).
Что бы объяснить появление особей с рекомбинантными признаками, необходимо вспомнить деление мейоза.
А именно то что в профазу первого мейоза гомологичные хромосомы конъюгируют (сближаются) и могут обмениваться гомологичными участками. Этот процесс называется кроссинговером. Он необходим для повышения разнообразия потомков.
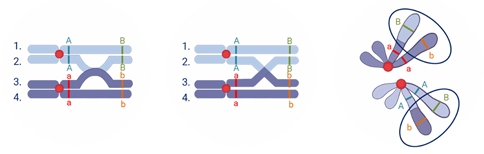
При кроссинговере происходит нарушение закона Моргана, и гены одной хромосомы не наследуются сцеплено так как часть из них заменяется на аллельные гены гомологичной хромосомы.
Происходит обмен участками хромосом между генами А и В, появляются гаметы А-большое бэ-малое и а-малое Бэ-большое, и, как следствие, в потомстве образуются четыре группы фенотипов.
Процесс кроссинговера был открыт Томосом Морганом и его сотрудниками при очередном скрещивании дрозофил.
Явление кроссинговера позволило лаборатории Томаса Моргана установить расположение каждого гена в хромосоме. И составить хромосомные карты.
Хромосомная карта — это графическое изображение хромосомы, на котором определённые локусы (гены) отмечены соответственно расстоянию между ними.
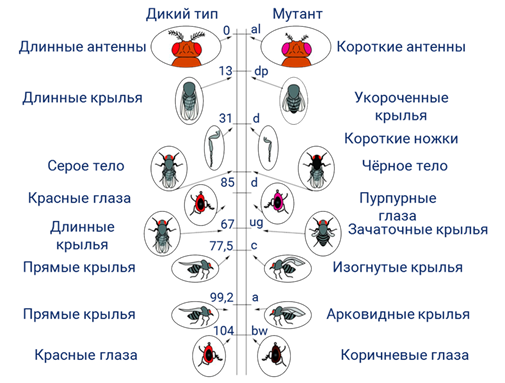
Определить в каких участках хромосом произойдёт кроссинговер достаточно сложно. Томас Морган доказал, что частота кроссинговера между генами прямо пропорциональна расстоянию между ними в хромосоме. Другими словами, можно сказать что, чем дальше гены находятся друг от друга в хромосоме, тем чаще между ними происходит кроссинговер.
Например, если рассматривать 2 гена А и D мы можем увидеть 2 случая. Если два гена А и D находятся по разные стороны перекрёста, то они разойдутся в разные хромосомы при любом варианте кроссинговера. В данном случае А-большое дэ-малое и а-малое D-большое.
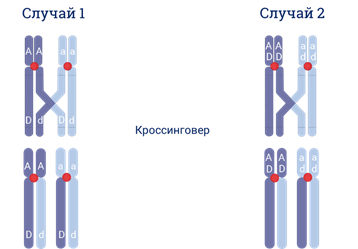
Но если гены, например, А-большое и D-большое расположены в хромосоме рядом…по одну сторону от перекрёста. Тогда после прохождения кроссинговера новых сочетаний аллелей данных двух генов мы не увидим.
Различают неполное и полное сцепление. Неполное сцепление — это разновидность сцепленного наследования, при котором гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Полное сцепление — это разновидность сцепленного наследования, при котором гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
Исходя из своих открытий Томас Морган сформулировал хромосомную теорию наследственности.
Первое положение этой теории гласит: ген представляет собой участок хромосомы. Хромосомы, таким образом представляют собой группу сцепления генов.
Второе положение утверждает, что, аллельные гены (гены, которые отвечают за один признак) расположены в строго определённых местах (локусах) гомологичных хромосом.
Согласно третьему положению, гены располагаются в хромосомах линейно, то есть друг за другом.
В ещё одном дополнительном положении говорится что в процессе образования гамет между гомологичными хромосомами происходит конъюгация, в результате которой они могут обмениваться аллельными генами, т.е. может происходит кроссинговер.
 0
0 12626
12626

