 Получите свидетельство
Получите свидетельство Вход
Вход



Организмы представляют собой сложную скоординированную систему, в которой все процессы взаимосвязаны. Связь процессов друг с другом в организме в значительной мере определяется взаимодействием генов между собой. Взаимоотношения между генами и признаками носят сложный характер.
Один и тот же ген может оказывать влияние на развитие нескольких признаков. А один и тот же признак может развиваться под влиянием многих генов.
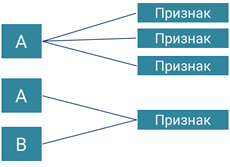
Различают две основные группы взаимодействия генов: взаимодействие между аллельными генами и взаимодействие между неаллельнимы генами.
На одном из уроков мы с вами уже рассмотрели взаимодействия между аллельными генами: полное доминирование, неполное доминирование и кодоминирование.
Взаимодействие неаллельных генов
К такому взаимодействию относят: дополнительное (комплементарное) взаимодействие, эпистаз и полимерное действие генов.
Однако следует понимать, что это не физическое взаимодействие самих генов, а взаимодействие белков, которые эти гены кодируют.

В цитоплазме происходит взаимодействие между белками – ферментами или между веществами, которые образовываются под влиянием этих ферментов.
Дополнительное (комплементарное) взаимодействие − это такой тип взаимодействия неаллельних генов, когда один доминантный ген дополняет действие другого неаллельного доминантного гена, и они вместе определяют новый признак, который отсутствует у родителей. Причём соответственный признак развивается только в присутствии обоих неаллельних генов.

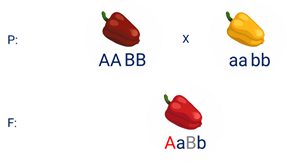
Например, при скрещивании двух чистых линий душистого горошка, которые имеют белые цветки у гибридов первого поколения все цветки будут пурпурными.

Почему так происходит? Дело в том, что в данном случае доминантные неаллельные гены А-большое и Бэ-большое, каждые в отдельности кодируют недостаточное количество пурпурного пигмента для окраски цветка.
И только при наличии обоих неаллельных доминантных генов А-большое и Бэ-большое, которые кодируют достаточное количество пигмента цветки окрашиваются в пурпурный цвет.
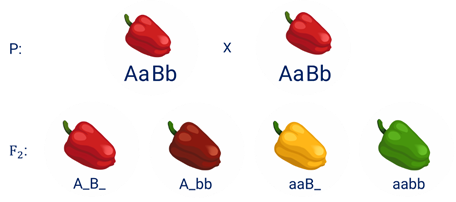
По принципу комплементарности наследуется окраска плодов у перца. Цвет плодов обеспечивается двумя типами пигментов – хлорофиллом и пигментами красного и жёлтого цвета.
В зависимости от их сочетания плоды могут иметь четыре различных варианта окраски.
Доминантный аллель А-большое контролирует образование красного пигмента, тогда как рецессивный аллель а-малое контролирует образование жёлтого пигмента. Доминантный аллель Бэ-большое блокирует синтез хлорофилла, не влияя на образование пигментов, тогда как рецессивный аллель бэ-малое не влияет на его синтез и такие растения синтезируют хлорофилл нормально.
При скрещивании растений с коричневыми плодами и растений с жёлтыми плодами в первом поколении все потомство будет иметь красные плоды.
А при скрещивании гетерозиготных растений перца из первого поколения. Во втором поколении будет наблюдаться расщепление.
Итак, дополнительным называют такой вид взаимодействия генов, когда для проявления признака необходимо присутствие неаллельных генов А и Бэ. Эти гены называют дополнительными или комплементарными.

Следующее взаимодействие неаллельных генов, которое мы рассмотрим это эпистаз.
Это вид взаимодействия неаллельных генов, при котором один ген подавляет (не даёт проявиться в фенотипе) другой ген.
Эпистаз противоположен комплементарному взаимодействию, здесь ген не дополняет действие другого, а подавляет.

Гены, которые подавляют действие других генов, называют генами-ингибиторами. Подавление могут вызывать как доминантные, так и рецессивные гены, и в зависимости от этого различают эпистаз доминантный и рецессивный.
Доминантный эпистаз характерен для окраски плодов у тыквы.
У этого растения известно три цвета плодов: белая, жёлтая и зелёная. Доминантный аллель гена А определяет жёлтую окраску тыквы, рецессивный аллель а-малое – зелёный цвет. Второй ген Бэ-большое проявляет эпистатическое действие – подавляет образование пигмента как жёлтого, так и зелёного цвета, делая их белыми. Рецессивный аллель бэ-малое – не влияет на проявление окраски плодов тыквы.
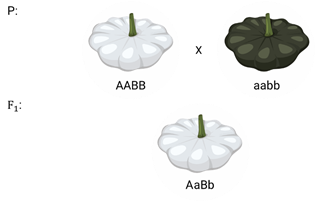
При скрещивании растений с белыми и зелёными плодами, все потомство первого поколения будет белым. Так как ген Бэ-большое подавляет действие гена А-большое.
А во втором поколении наблюдается расщепление признаков. В этом случае рецессивная гомозигота (a-малое a-малое бэ-малое бэ-малое) фенотипически отличима от гетерозиготных классов.
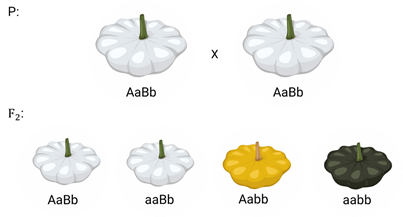
Таким образом растения, имеющие в своём генотипе хотя бы один доминантный ген Бэ-большое, будут образовывать белые плоды независимо от того, какие аллели гена А-большое будут присутствовать в их генотипах.
Следующий тип взаимодействия неаллельных генов — полимерное действие генов.
Полимерия ─ это вид взаимодействия двух и более неаллельных генов, доминантные аллели которых однозначно влияют на развитие одного и того же признака.
Часто признаки в организмах могут быть выражены слабее или сильнее – например вес, удойность, плодовитость. Такие признаки называют количественными, они определяются несколькими генами.

Полимерные гены обозначаются одной буквой, аллели одного локуса имеют одинаковый цифровой индекс.
Действие признаков суммируется, и чем больше в генотипе доминантных пар генов, которые влияют на количественный признак, тем сильнее он проявляются.
Полимерное действие генов может быть кумулятивным и некумулятивным.
При кумулятивной полимерии интенсивность значения признака зависит от суммирующего действия генов: чем больше доминантных аллелей, тем больше степень выраженности признака.
При некумулятивной полимерии количество доминантных аллелей на степень выраженности признака не влияет, а признак проявляется при наличии хотя бы одного из доминантных аллелей.
Приведём пример кумулятивной полимерии.
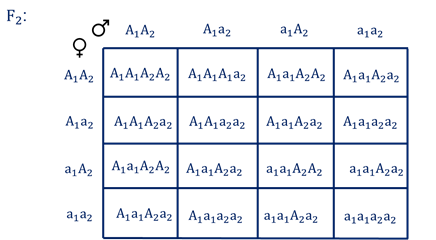
Бурый цвет зерна пшеницы обусловлен доминантными генами из двух пар аллелей – А-большое 1 и А-большое 2.
Поэтому самые яркие семена имеют растения, у которых четыре доминантных гена А1 А1 А 2 А2. У растений с генотипом А1 а1 а2 а2 или а1 а1 А2 а2 – зерно окрашено слабо. А у растений с генотипом а1 а1 а2 а2 − зерно не окрашено.
Например, при скрещивании растения с ярко бурыми семенами и растения с неокрашенными семенами в первом поколении все организмы получаются со слабо окрашенными семенами, так как им досталась половина доминантных генов от первого родителя.
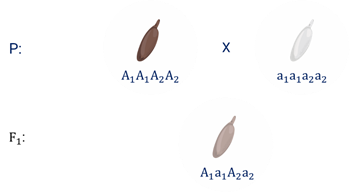
При скрещивании гетерозиготных растений из первого поколения происходит вот такое расщепление.
Другой пример полимерного действия генов – наследование окраски кожи человека. Цвет кожи определяется четырьмя генами, расположенными в четырёх различных хромосомах. Эти гены отвечают за синтез тёмного пигмента кожи – меланина.
У европейцев чаще всего встречаются рецессивные гены, отвечающие за синтез меланина. А у самых темных африканцев доминантные.

Итак, мы рассмотрели типы взаимодействия неаллельных генов.
При комплементарном взаимодействии один ген дополняет действие другого гена.
При эпистазе один ген подавляет действие другого.
При полимерном взаимодействии проявление признака зависит от двух и более пар неаллельных генов.
Бывает и так, что несколько признаков зависит от одного гена. Такое множественное действие гена называется плейотропностью.
Плейотропное действие генов впервые было обнаружено ещё Грегором Менделем, который заметил, что у растений с пурпурными цветками стебли всегда темнее, чем у растений с белыми цветками. Развитие этих признаков определяется действием одного гена.
Плеотропное действие генов встречается и у человека. Наследственная болезнь — серповидно-клеточная анемия проявляется в результате замены одной аминокислоты в молекуле гемоглобина, что приводит к изменению формы эритроцитов.
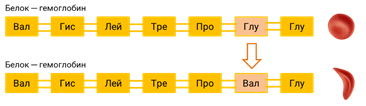
Одновременно с этим возникают нарушения в сердечно-сосудистой, нервной, пищеварительной и выделительной системах. Это приводит к тому, что гомозиготный по этому заболеванию ребёнок погибает в детстве.
Уже давно генетики встречались с многочисленными фактами, которые не укладывались в рамки хромосомной теории наследственности.
Мы говорили, что в генах закодирована информация об определённых белках организма и эти определённые белки, синтезируясь в клетках, и являются уже основой проявления того или иного признака организма.
Вы знаете, что в наследовании признаков ведущую роль отводят ДНК, которая находится в ядре.
Однако, существует путь передачи наследственной информации через цитоплазму клетки. Такой вид наследственности называют цитоплазматической или нехромосомной.
Цитоплазматическая наследственность – это наследственность, которая осуществляется с помощью молекул ДНК, находящихся в пластидах и митохондриях. В этом случае признаки не наследуются по законам Менделя, так как указанные молекулы ДНК находятся вне хромосом.
При изучении митохондрий и пластид мы говорили они имеют собственную ДНК.
В генах этих ДНК закодирована информация о белках. А значит ДНК пластид и митохондрий несут какую-либо информацию. Благодаря ДНК митохондрии и пластиды способны самовоспроизводится.
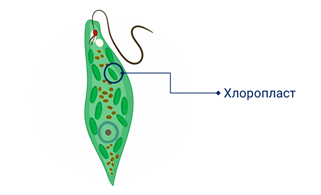
Если клетка эвглены зелёной утрачивает пластиды, то образовать их снова она уже не может. Так, как и ДНК которая в ней находилась тоже разрушилась. И даже несмотря на то что сохранилось ядро пластиды не образуются. Так как информация хранилась в самих пластидах.
Вы знаете, что на свету эвглена зелёная питается автотрофно. Автотрофному питанию способствуют хлоропласты. В темноте они естественно не функционируют, так как нет света. Поэтому эвглена переходит на гетеротрофные питание.

Если же эвглену долгое время выдерживать в темноте, то её пластиды перестают делится. А при размножении (делении) самой эвглены, появляются особи, которые по этой причине не получили хлоропласты. То есть появляются бесхлоропластные особи.

Итак, мы сказали, что в ДНК митохондрий и пластид закодирована информация. Это информация может передаваться по наследству через цитоплазму клетки. В таком случае говорят о цитоплазматической наследственности.
При оогамной форме полового процесса, где женская гамета представлена яйцеклеткой, а мужская сперматозоидом, признаки, которые наследуются цитоплазматически, передаются только через яйцеклетку. То есть по материнской линии.
Почему? Если вспомнить строение сперматозоидов и яйцеклеток. А также то как происходит процесс оплодотворения. То все становиться сразу ясно. В цитоплазме яйцеклеток митохондрий может быть много, а вот в цитоплазме мужских гамет этих органоидов обычно нет.
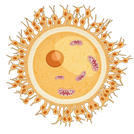
Митохондрии находятся в шейки сперматозоида. При оплодотворении в яйцеклетку они не попадают. Вспомните проникает лишь генетический материал сперматозоида. Таким образом, все митохондрии и пластиды зиготы достаются ей в наследство только от материнского организма. И все признаки которые несут их ДНК, передаются от матери.
Например, пестролистность растения львиный зев передаётся по материнской линии, так как яйцеклетка содержит хлоропласты в виде пропластид.

Пластиды размножаются делением, но поскольку не все они содержат хлорофилл (в силу мутаций), растение получается пестролистным.
 0
0 9869
9869

