 Получите свидетельство
Получите свидетельство Вход
Вход



Размножение – это одно из обязательных свойств любого живого организма. Оно заключается в увеличении числа особей.
Различают бесполое и половое размножение растений.
Бесполое размножение подразделяют на спорообразование и вегетативное.
Половое размножение происходит при помощи особых половых клеток –гамет.
При бесполом размножении быстро увеличивается численность вида, все потомки имеют абсолютно такой же генотип, что и родительская особь. А также не происходит увеличения генетического разнообразия, которое может оказаться очень полезным при изменении условий существования вида.
По этой причине большинство живых организмов на Земле размножаются половым путём.
Сущность полового размножения заключается в слиянии генетической информации родителей, благодаря чему генетическое разнообразие в потомстве увеличивается.
У покрытосеменных растений половое размножение связано с цветком.
Важные части цветка – это пестик и тычинка. С их участием происходят сложные процессы полового размножения –опыление и оплодотворение.
Но сперва в будущем цветке начинают образовываться половые клетки.
В жизненном цикле цветковых растений наблюдается смена поколений.
У растений выделяют диплоидное поколение – бесполое, или спорофит, и гаплоидное поколение – половое, или гаметофит.
Гаметы образуются в результате митоза, а споры – в результате мейоза. И гаметы, и споры образуются в цветке, поэтому цветок является органом и бесполого, и полового размножения.
Пыльцевое зерно является спорой (микроспорой), а не мужской гаметой, так как в нем самом развиваются мужские гаметы.
У цветковых растений мужские гаметофиты столь малы, что помещаются внутри оболочки пыльцевого зерна и состоят всего лишь из нескольких клеток.
Женский гаметофит цветковых (зародышевый мешок) помещается внутри семяпочки и состоит в наиболее распространённом случае из 7 клеток (содержит 8 – либо 7 после слияния двух ядер в центральной клетке и образования вторичного ядра).
Посмотрим, как образуется мужской гаметофит.
Итак, Микроспорогенез
В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делений становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
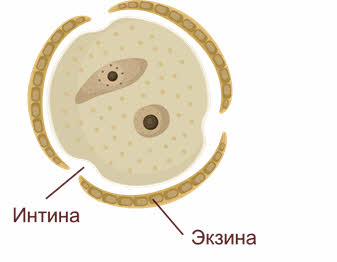
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, поверхность её либо гладкая, либо шероховатая, приспособленная для переноса пыльцы и прилипания её к рыльцу пестика.
Этим заканчивается микроспорогенез, вслед за образованием одноядерной микроспоры начинается микрогаметогенез.
Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и её ядро не делятся. В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться ещё в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, называются спермиоклетками, или спермиями.
Таким образом, из одной споры (микроспоры) с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: два из них – спермии и одно – вегетативное. При образовании пыльцевой трубки это вегетативное ядро переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены российским и советским цитологом и эмбриологом растений Сергеем Гавриловичем Навашиным в 1910 г. на лилейных растениях.
После образования гамет пыльник созревает, и пыльца высыпается. Она несёт только генетическую информацию.
Посмотрим, как происходит мегаспорогенез и мегагаметогенез цветковых.
У покрытосеменных растений женский гаметофит – это зародышевый мешок, который закладывается и развивается внутри семяпочки.
Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез.
В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка археспория растёт, превращаясь в материнскую клетку мегаспоры.
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной. Однако только одна из них продолжает развиваться, остальные три дегенерируют, судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем её ядро претерпевает ряд делений. При этом сама клетка не делится, а делится только ядро.
У разных систематических групп растений число делений ядра мегаспоры может варьировать от одного до трёх. У большинства растений (70 % видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других – в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
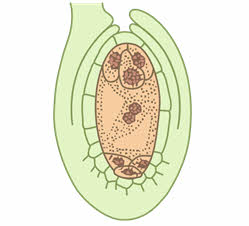
В дальнейшем от каждой из двух полярных четвёрки ядер к центру отходит по одному ядру, которые сливаются, образуя вторичное (центральное) ядро зародышевого мешка. Затем цитоплазма обособляется вокруг ядер гаметофита, который из ядерной стадии развития переходит в клеточную. Три ядра, оставшиеся вблизи халазального полюса, преобразуются в три клетки (антиподы), которые питают гаметофит. Три ядра вблизи микропиле отделяются клеточными перегородками, образуя отдельные клетки: крупную центральную яйцеклетку и две боковые клетки синергиды. Вся цитоплазма, расположенная между антиподами с одной стороны и клетками яйцевого комплекса с другой (яйцеклетка и две синергиды), называется центральной клеткой. В ней находится диплоидное вторичное (центральное) ядро.
На этом этапе женский гаметофит уже полностью сформирован и состоит из шести гаплоидных клеток (одной яйцеклетки, двух синергид и трёх антипод) и одной диплоидной (центральной). Его строение внешне напоминает мешочек, поэтому женский гаметофит покрытосеменных называется зародышевым мешком.
Пылинка попадает на рыльце пестика, и происходит опыление.
Опыление – это перенос пыльцевых зёрен на рыльце пестика, у голосеменных пыльцевые зерна при опылении попадают непосредственно на семязачаток.
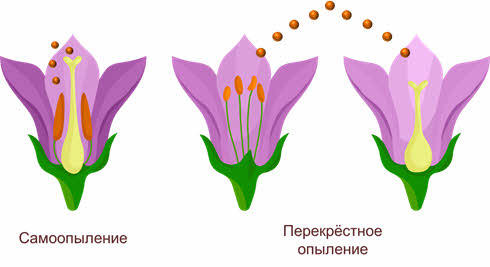
Имеется два основных типа опыления: самоопыление (автогамия) (когда растение опыляется собственной пыльцой) и перекрёстное опыление (аллогамия).
При самоопылении исключён обмен генетической информацией, поскольку пыльцевые зерна попадают на рыльце пестика либо с одной из тычинок этого же цветка, либо с другого цветка, расположенного на том же растении. Это приводит к появлению чистых линий гомозиготных популяций в пределах одного вида, неспособных обмениваться мутировавшими генами, поэтому процессы видообразования в этих популяциях идут самостоятельно.
Перекрёстное опыление – это перенос пыльцы одного растения на рыльце другого. Этот тип опыления встречается более часто, чем самоопыление, между разными особями одного вида происходит обмен аллелями, что приводит к увеличению доли гетерозиготных организмов.
Безусловно, перекрёстное самоопыление имеет большие преимущества по сравнению с самоопылением, поскольку возникшие мутации свободно распространяются в пределах популяции.
Способы опыления у перекрёстноопыляемых цветковых растений весьма разнообразны. Их можно разделить на две группы. Первая: перенос пыльцы осуществляется главным образом насекомыми, а также некоторыми позвоночными (птицами и летучими мышами). Соответственно, различают энтомофилию, орнитофилию и зоофилию. Растения, опыляемые животными, обычно имеют яркоокрашенные крупные цветки. Мелкие цветки, как правило, собраны в соцветия, что зрительно их увеличивает. Для привлечения опылителей служит нектар или большое количество пыльцы, которую опылители охотно поедают.
Вторая группа: пыльца переносится абиотическими факторами – ветром и реже водой, в связи с чем различают анемофилию и гидрофилию.
Оплодотворение.
После попадания пыльцы на рыльце начинается прорастание пыльцевого зерна.
Экзина мужского гаметофита прорывается в области борозды или поры прорастания, и начинает формироваться пыльцевая трубка, которая растёт, проникает в рыльце, через столбик движется по направлению к завязи, где находится семязачаток.
Трубка растёт на кончике. В растущую пыльцевую трубку из пыльцевого зерна перемещается ядро клетки-трубки, а также спермин.
Обычно развитие пыльцевой трубки происходит при уже сформированных семязачатках в завязи. Достигнув завязи, пыльцевая трубка через микропиле проникает внутрь одного из находящихся там семязачатков.
Там пыльцевая трубка направляется к яйцевому аппарату, проникает в одну из синергид и освобождает спермин. Один из спермиев сливается с яйцеклеткой, в результате чего образуется зигота.
Другой спермий сливается с полярными ядрами центральной клетки, образуя триплоидное ядро (с тройным набором хромосом). Зигота даёт начало зародышу, а из триплоидной центральной клетки образуется ткань эндосперма.
Таким образом оплодотворение, при котором одна мужская гамета сливается с яйцеклеткой, а вторая ― с вторичным ядром, называется двойным оплодотворением.
Механизм оплодотворения цветковых –двойное оплодотворение, открыто отечественным учёным Сергеем Гавриловичем Навашиным в 1898 году.
Таким образом, оплодотворённый семязачаток развивается в семя, из зиготы возникает зародыш, из триплоидной центральной клетки – эндосперм, а из внешней части семязачатка (интегументов) образуется семенная кожура, из стенок завязи цветка – стенки плода.
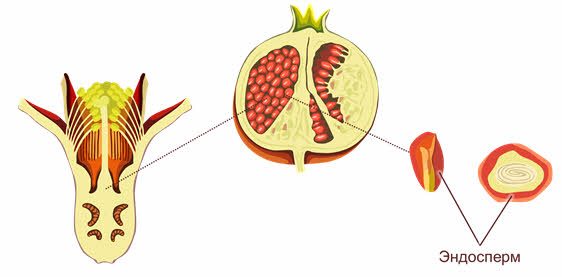
Развитие эндосперма.
Эндосперм цветковых полностью отличается от первичного эндосперма голосеменных. У них первичный эндосперм представляет собой гаплоидную вегетативную ткань женского гаметофита, где накапливаются питательные вещества семени.
Эндосперм у покрытосеменных происходит из триплоидной центральной клетки, в образовании которой участвуют вторичное ядро женского гаметофита и ядро спермия.
У одних цветковых (например, злаков) эндосперм сильно разрастается и занимает большую часть семени, оттесняя на периферию маленький зародыш.
У других (к пример у бобовых) весь эндосперм поглощается зародышем и используется его семядолями, которые становятся самыми большими структурами зрелого семени.

 13231
13231

