
УГЛЕВОДЫ
УГЛЕВОДЫ – это альдегидо- и кетопроизводные
многоатомных спиртов
R
R
Функции углеводов :
- энергетическая (моносахариды, гомополисахариды)
- структурная (гетерополисахариды)
- углеводы входят в состав сложных белков – гликопротеинов ( ферментов, гормонов,
рецепторов, иммуноглобулинов и т.д.)
- углеводы используются в синтезе нуклеотидов (в синтезе ДНК, РНК, коферментов)
кетоза
альдоза
Моносахариды:
Дисахариды:
Полисахариды:
- мальтоза
- сахароза
- лактоза
- крахмал (амилоза и амилопектин)
- гликоген
- целлюлоза
- глюкоза
- фруктоза
- галактоза
фруктоза
галактоза
глюкоза
Крахмал – разветвленный гомополисахарид, состоит из мономера глюкозы, Мономеры линейных участков (амилозы) соединены 1,4-гликозидными связями, а в местах разветвления – 1,6-гликозидными связями.
1,6-гликозидная
связь
Гликоген – полисахарид, по структуре подобный крахмалу, но более разветвленный (т.е. содержит больше 1,6-гликозидных связей, а значит большее количество концевых мономеров, что облегчает его распад ).
1,4-гликозидная
связь
Основные процессы в обмене углеводов
1. Переваривание углеводов
2. Синтез гликогена - гликогенез
3. Распад гликогена - гликогенолиз.
4. Дихотомический распад глюкозы - непрямое окисление глюкозы при делении ее молекулы на две триозы
4.1. Анаэробный гликолиз - окисление глюкозы в безкислородных условиях с образованием двух молекул лактата и 2 АТФ (за счет субстратного фосфорилирования). Происходит в активно работающей мышце, эритроцитах и др.
4.2. Аэробный гликолиз - окисление глюкозы в кислородных условиях с образованием двух молекул пирувата и 8 АТФ (2 - за счет субстратного фосфорилирования и 6 - за счет окислительного фосфорилирования).
Далее пируват в этих условиях окисляется в ацетил-КоА с образованием 3АТФ (за счет окислительного фосфорилирования), а ацетил-КоА сгорает в ЦТК с образованием 12АТФ, т.е. при окислении пирувата до углекислого газа и воды образуется 15АТФ, а т.к. из глюкозы обазуется 2 пирувата, то 30АТФ. Всего полное сгорание глюкозы в кислородных условиях дает 30+8=38АТФ.
4.3. Спиртовое брожение - превращение глюкозы в этиловый спирт под действием микроорганизмов (дрожжей).
5. Глюконеогенез - синтез глюкозы из неуглеводов: аминокислот (белков), глицерина (липидов), пирувата, лактата. Является процессом, обратным гликолизу, за исключением необратимых реакций гликолиза, которые в глюконеогенезе идут с помощью других ферментов: глюкозо-6-фосфатазы, фруетозо-1,6-бисфосфатазы,фосфоенолпируваткарбоксики-назы (ФЕПКК), пируваткарбоксилазы, а также различных трансаминаз.
Происходит в печени, активируется при голодании под действием глюкокортикоидов
6. Апотомический распад глюкозы - прямое окисление глюкозы - пентозо-фосфатный путь.
Глюкоза окисляется и декарбоксилируется (отщепление apex - верхушки - одого атома углерода в виде углекислого газа) с образованием фосфопентоз и НАДФН.
Фосфопентозы используются в синтезе нуклеотидов, НАДФН - в синтезе жиров
7. Взаимопревращения моносахаров
Переваривание углеводов
Переваривание углеводов – это гидролиз олиго- и полисахаридов, т.е. расщепление гликозидных связей под действием воды с помощью гидролитических ферментов.
Ротовая полость
Начинается переваривание углеводов в ротовой полости, гле действует -амилаза слюны (рН opt = 6 , 7 ), катализирующая гидролиз 1,4-гликозидных связей в молекуле крахмала. Этот фермент является эндоамилазой , т.е. расщепляет связи, удаленные от конца цепи. В результате образуются олигосахариды (декстрины) и небольшое количество дисахарида мальтозы.
-амилаза
слюны
1,6-
гликозидаза
В желудке (рН=1,5-2) переваривание углеводов
не происходит
Тонкий кишечник
панкреатическая
-амилаза
В верхнем отделе тонкой кишки действует панкреатическая -амилаза, которая гидролизует 1,4-гликозидные связи, последовательно отщепляя молекулы мальтозы от декстринов. 1,6-гликозидные связи в декстринах гидролизуют 1,6-гликозидазы.
На дисахариды в кишечнике действуют соответствующие ферменты ( мальтаза, сахараза и лактаза), расщепляющие их до моносахаридов:
глюкоза + глюкоза
+
мальтаза
+
глюкоза + фруктоза
сахараза
+
глюкоза + галактоза
лактаза
Превращение глюкозы в тканях
Первая реакция превращения глюкозы в тканях – активация глюкозы путем фосфорилиро-вания с участием АТФ. Катализирует ее во многих тканях гексокиназа , а в клетках печени – глюкокиназа . Глюкокиназа специфична для глюкозы и работает при высоких ее концентрациях ( К M = 12ммоль/л), а гексокиназа фосфорилирует разные гексозы и ингибируется глюкозо-6-фосфатом ( К M = 0,1ммоль/л).
АТФ
АДФ
гексокиназа
глюкокиназа
глюкоза-6-фосфат
глюкоза
Эта реакция «запирает» глюкозу в клетке, т.к. клеточная мембрана непроницаема для фосфорилированной глюкозы.
Синтез гликогена
Гликоген – животный гомополисахарид, резервная форма глюкозы , в основном депонируется в печени и скелетных мышцах . Гликоген животных, как и амилопектин растений, представляет собой разветвленный полимер глюкозы , в котором остатки глюкозы соединены α(1→4)-гликозидной связью. Связи в точках ветвления находятся в положении α(1→6) примерно каждого 10-го остатка.
ФФ
УТФ
фосфо-
глюкомутаза
УДФ-глюкозо-
пирофосфорилаза
глюкоза-6-фосфат
глюкозо-1-фосфат
УДФ-глюкоза
Хранение в организме собственно глюкозы неприемлемо из-за ее высокой растворимости: высокие концентрации глюкозы создают в клетке высоко гипертоническую среду, что приводит к притоку воды. Напротив, нерастворимый гликоген осмотически почти неактивен и компактен.
Гранулы гликогена
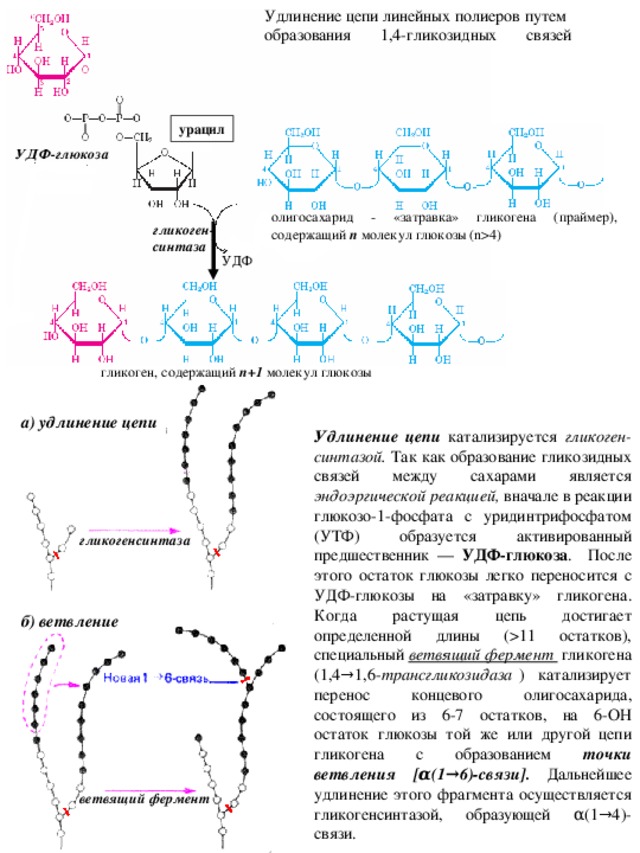 4) гликоген- синтаза УДФ гликоген, содержащий n +1 молекул глюкозы а) удлинение цепи Удлинение цепи катализируется гликоген-синтазой. Так как образование гликозидных связей между сахарами является эндоэргической реакцией, вначале в реакции глюкозо-1-фосфата с уридинтрифосфатом (УТФ) образуется активированный предшественник — УДФ-глюкоза . После этого остаток глюкозы легко переносится с УДФ-глюкозы на «затравку» гликогена. Когда растущая цепь достигает определенной длины (11 остатков), специальный ветвящий фермент гликогена (1,4→1,6- трансгликозидаза ) катализирует перенос концевого олигосахарида, состоящего из 6-7 остатков, на 6-ОН остаток глюкозы той же или другой цепи гликогена с образованием точки ветвления [α(1→6)-связи]. Дальнейшее удлинение этого фрагмента осуществляется гликогенсинтазой, образующей α(1→4)-связи. гликогенсинтаза б) ветвление ветвящий фермент " width="640"
4) гликоген- синтаза УДФ гликоген, содержащий n +1 молекул глюкозы а) удлинение цепи Удлинение цепи катализируется гликоген-синтазой. Так как образование гликозидных связей между сахарами является эндоэргической реакцией, вначале в реакции глюкозо-1-фосфата с уридинтрифосфатом (УТФ) образуется активированный предшественник — УДФ-глюкоза . После этого остаток глюкозы легко переносится с УДФ-глюкозы на «затравку» гликогена. Когда растущая цепь достигает определенной длины (11 остатков), специальный ветвящий фермент гликогена (1,4→1,6- трансгликозидаза ) катализирует перенос концевого олигосахарида, состоящего из 6-7 остатков, на 6-ОН остаток глюкозы той же или другой цепи гликогена с образованием точки ветвления [α(1→6)-связи]. Дальнейшее удлинение этого фрагмента осуществляется гликогенсинтазой, образующей α(1→4)-связи. гликогенсинтаза б) ветвление ветвящий фермент " width="640"
Удлинение цепи линейных полиеров путем
образования 1,4-гликозидных связей
урацил
УДФ-глюкоза
олигосахарид - «затравка» гликогена (праймер), содержащий n молекул глюкозы ( n 4)
гликоген-
синтаза
УДФ
гликоген, содержащий n +1 молекул глюкозы
а) удлинение цепи
Удлинение цепи катализируется гликоген-синтазой. Так как образование гликозидных связей между сахарами является эндоэргической реакцией, вначале в реакции глюкозо-1-фосфата с уридинтрифосфатом (УТФ) образуется активированный предшественник — УДФ-глюкоза . После этого остаток глюкозы легко переносится с УДФ-глюкозы на «затравку» гликогена. Когда растущая цепь достигает определенной длины (11 остатков), специальный ветвящий фермент гликогена (1,4→1,6- трансгликозидаза ) катализирует перенос концевого олигосахарида, состоящего из 6-7 остатков, на 6-ОН остаток глюкозы той же или другой цепи гликогена с образованием точки ветвления [α(1→6)-связи]. Дальнейшее удлинение этого фрагмента осуществляется гликогенсинтазой, образующей α(1→4)-связи.
гликогенсинтаза
б) ветвление
ветвящий фермент
Распад гликогена в тканях
Гликоген печени никогда не расщепляется полностью. Как правило, укорачиваются только невосстанавливающие концы древовидной структуры путем фосфоролиза, т.е. присоединения фосфорной кислоты по месту разрыва связи.
Н 3 РО 4
Гликоген ( n молекул глюкозы)
гликоген-
фосфорилаза
Гликогенфосфорилаза отщепляет от неразветленных цепей гликогена по одному мономеру в виде глюкозо-1-фосфата
Гликоген ( n -1 молекул глюкозы)
Глюкозо-1-фосфат
1,6-связь
Гликогенфосфорилаза катализирует фосфоролиз 1,4-связей, т.е. расщепление линейных полимеров. Разрыв 1,6-связей катализирует деветвящий фермент , который обдадает:
- гликозилтрансферазной активностью , т.е. переносит олигосахарид из трех глюкоз в конец неразветвленной цепи, при этом остается одна молекула глюкозы , связанная 1,6-связью;
- 1,6-гликозидазной активностью, т.е. расщепляет 1,6-связи с образованием свободной глюкозы и линейного полимера , на который далее действует гликогенфосфорилаза, отщепляя по одной молекуле глюкозо-1-фосфата.
гликоген
гликоген-
фосфорилаза
молекулы
глюкозо-1-фосфата
деветвящий
фермент
гликозилтрансферазная
активность
деветвящий
фермент
1,6-гликозидазная
активность
глюкоза
линейный (1,4-) полимер - субстрат для дальнейшего действия гликогенфосфорилазы

Регуляция гликогенолиза и гликогенеза
(синтеза и распада гликогена)
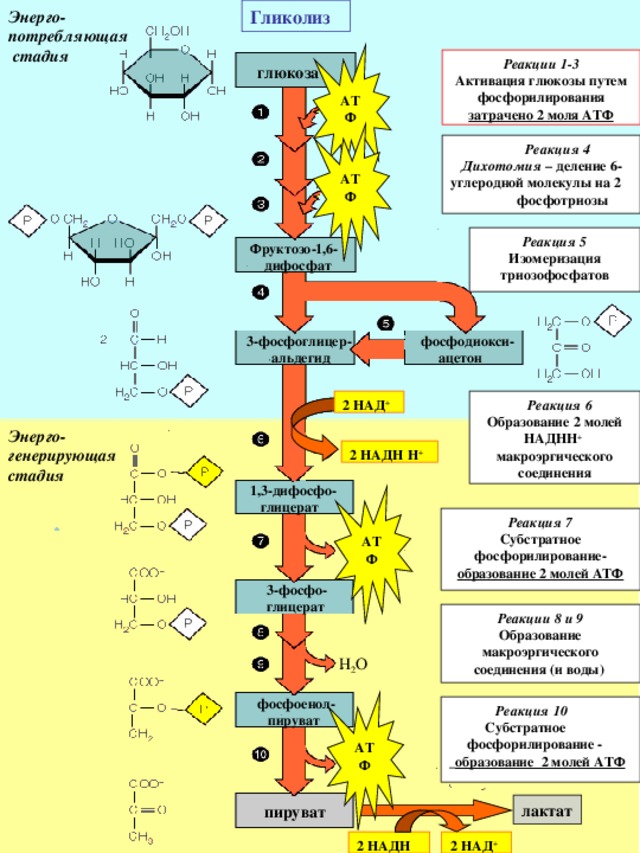
Гликолиз
Энерго-
потребляющая
стадия
АТФ
Реакции 1-3
Активация глюкозы путем
фосфорилирования
затрачено 2 моля АТФ
глюкоза
АТФ
Реакция 4
Дихотомия – деление 6-углеродной молекулы на 2 фосфотриозы
Реакция 5
Изомеризация
триозофосфатов
Фруктозо-1,6-
дифосфат
фосфодиокси-
ацетон
3-фосфоглицер-
альдегид
2 НАД +
Реакция 6
Образование 2 молей НАДНН + макроэргического соединения
Энерго-
генерирующая
стадия
2 НАДН Н +
1,3-дифосфо-
глицерат
АТФ
Реакция 7
Субстратное
фосфорилирование-
образование 2 молей АТФ
3-фосфо-
глицерат
Реакции 8 и 9
Образование макроэргического соединения (и воды)
Н 2 О
фосфоенол-
пируват
АТФ
Реакция 10
Субстратное
фосфорилирование -
образование 2 молей АТФ
лактат
пируват
2 НАД +
2 НАДН
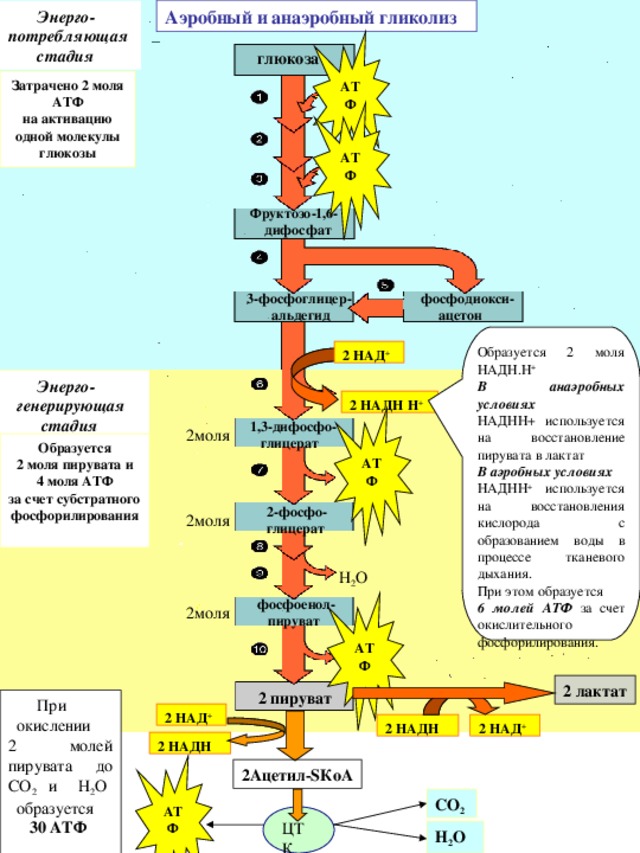
Аэробный и анаэробный гликолиз
Энерго-
потребляющая
стадия
АТФ
глюкоза
Затрачено 2 моля АТФ
на активацию
одной молекулы глюкозы
АТФ
Фруктозо-1,6-
дифосфат
фосфодиокси-
ацетон
3-фосфоглицер-
альдегид
Образуется 2 моля НАДН.Н +
В анаэробных условиях
НАДНН+ используется на восстановление пирувата в лактат
В аэробных условиях
НАДНН + используется на восстановления кислорода с образованием воды в процессе тканевого дыхания.
При этом образуется
6 молей АТФ за счет окислительного фосфорилирования.
2 НАД +
Энерго-
генерирующая
стадия
2 НАДН Н +
АТФ
1,3-дифосфо-
глицерат
2моля
Образуется
2 моля пирувата и
4 моля АТФ
за счет субстратного фосфорилирования
2-фосфо-
глицерат
2моля
Н 2 О
фосфоенол-
пируват
АТФ
2моля
2 лактат
2 пируват
При
окислении
2 молей пирувата до СО 2 и Н 2 О
образуется
30 АТФ
2 НАД +
2 НАД +
2 НАДН
2 НАДН
АТФ
2Ацетил- S КоА
СО 2
ЦТК
Н 2 О
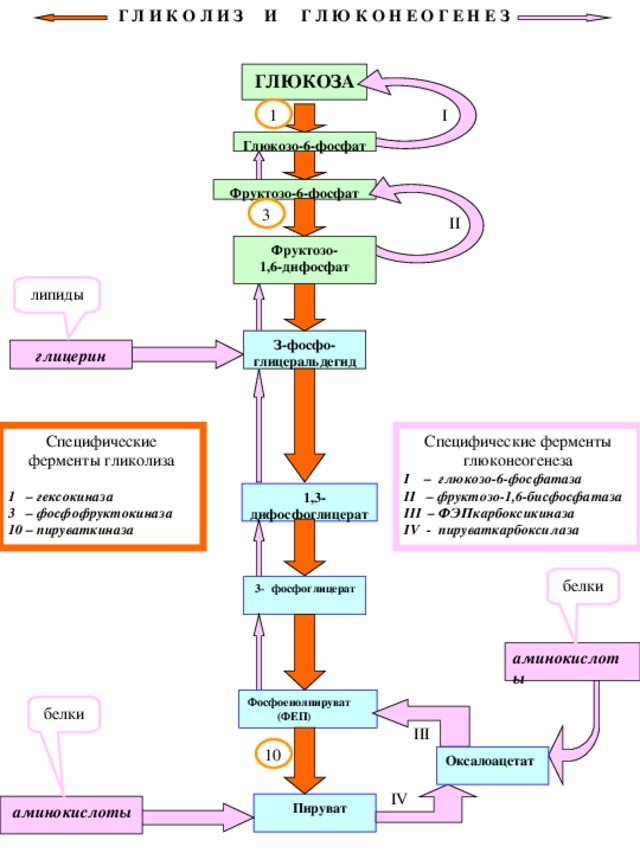
Г Л И К О Л И З И Г Л Ю К О Н Е О Г Е Н Е З
ГЛЮКОЗА
1
I
Глюкозо-6-фосфат
Фруктозо-6-фосфат
3
II
Фруктозо-
1,6-дифосфат
липиды
З-фосфо-
глицеральдегид
глицерин
Специфические ферменты глюконеогенеза
I – глюкозо-6-фосфатаза
II – фруктозо-1,6-бисфосфатаза
III – ФЭПкарбоксикиназа
IV - пируваткарбоксилаза
Специфические ферменты гликолиза
1 – гексокиназа
3 – фосфофруктокиназа
10 – пируваткиназа
1,3-дифосфоглицерат
белки
3- фосфоглицерат
аминокислоты
Фосфоенолпируват
(ФЕП)
белки
III
10
Оксалоацетат
IV
Пируват
аминокислоты

 Получите свидетельство
Получите свидетельство Вход
Вход












 Презентация по химии "Углеводы: основные представители класса" (0.4 MB)
Презентация по химии "Углеводы: основные представители класса" (0.4 MB)
 0
0 1076
1076 37
37 Нравится
0
Нравится
0



