 Получите свидетельство
Получите свидетельство Вход
Вход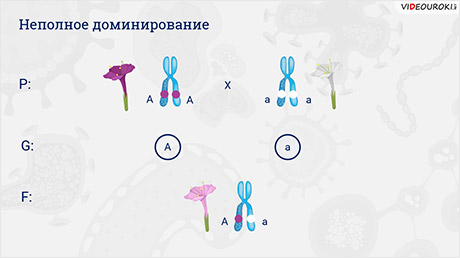



Организмы представляют собой сложную скоординированную систему, в которой все процессы взаимосвязаны. Связь процессов друг с другом в организме в значительной мере определяется взаимодействием генов между собой. Взаимоотношения между генами и признаками носят сложный характер.
Один и тот же ген может оказывать влияние на развитие нескольких признаков. А один и тот же признак может развиваться под влиянием многих генов.
Различают две основные группы взаимодействия генов: взаимодействие между аллельными генами и взаимодействие между неаллельными генами.

К взаимодействию между аллельными генами относят: полное доминирование, неполное доминирование и кодоминирование.
К взаимодействию между неаллельными генами относят: дополнительное (комплементарное) взаимодействие, эпистаз и полимерное взаимодействие генов.
Однако следует понимать, что это не физическое взаимодействие самих генов, а взаимодействие белков, которые эти гены кодируют.
В цитоплазме происходит взаимодействие между белками (ферментами) или между веществами, которые образутся под влиянием этих ферментов.
Гены, ответственные за развитие только одного признака (например, цвет семян), получили название аллельных генов. Аллели определяют противоположные варианты развития одного и того же признака. Обычно в качестве обозначения аллеля применяют сокращение названия соответствующего гена до одной или нескольких букв.
У диплоидных организмов, то есть у организмов, имеющих только две гомологичные хромосомы, по два аллеля каждого гена, поэтому мы можем обозначить их только двумя одинаковыми буквами или двумя разными.
Вы знаете, что доминантные признаки принято обозначать прописными латинскими буквами, а рецессивные — строчными. Что значит доминантный, а что значит рецессивный ген?
С ДНК участка гена синтезируется некая РНК с которой синтезируется белок.
Белок может быть функциональным, то есть катализировать какую-то реакцию.
Так же может синтезироваться и нефункциональный белок. В таком случае белок, который затем преобразуется в фермент, не будет катализировать никакую реакцию, и мы не увидим конечного продукта.
Рассмотрим взаимодействие между аллельными генами ― полное доминирование.
При полном доминировании одна из гетерозигот неотличима фенотипически от гомозигот. То есть в фенотипе гетерозигот присутствует продукт доминантного гена.
В каком случае это может происходить? Например, если ген кодирует фермент. В данном примере гомозигота несущая в себе два аллеля А-большое будет фенотипически красного цвета. Гетерозигота будет все равно красного цвета, так как функциолнальный аллель А-большое будет в полной мере обеспечивать организм белковым продуктом.
А гомозигота несущая два аллеля а-малое будет белого цвета потому что красного пигмента образовывать не будет.
Полное доминирование наблюдается при наследовании резус-фактора у человека. Вспомним, что резус-фактор — это антиген (белок), который находится на поверхности эритроцитов — красных кровяных телец.
Наличие резус-фактора эритроцитов обусловливается доминантным геном Rh (эр аш).
Генотип резус-положительного человека может быть двух видов: или Эр-большое аш-малое Эр-большое аш-малое (RhRh), или Эр-большое аш-малое эр-малое аш-малое (Rhrh); а генотип резус-отрицательного человека только эр-малое аш-малое эр-малое аш-малое (rhrh).
Если, например, мать — резус-отрицательная, а отец резус-положительный и гетерозиготен по этому признаку, то при данном типе брака с одинаковой вероятностью может родиться как резус-положительный, так и резус-отрицательный ребёнок.
Следующий тип взаимодействия не аллельных генов ― неполное доминирование. У травянистого растения ночной красавицы могут формироваться цветки разной окраски. Она бывает розовой, а её оттенки меняются от лососёвого до малинового, но может быть белой или жёлтой.
Если скрестить растение ночной красавицы, в хромосомах которого находятся доминантные гены (А-большое А-большое), отвечающие за пурпурный окрас цветков, с растением, в хромосомах которого находятся рецессивные гены (а-малое, а-малое), отвечающие за белый окрас цветков, то в первом поколении все цветки станут розового окраса. Почему?
Вы знаете, что гены кодируют белки, и часто это белки-ферменты. У одного родительского растения оба аллеля (A-большое A-большое) кодируют ферменты, ответственные за синтез пурпурного пигмента. Ферментов вырабатывается много, и цвет цветков пурпурный. У второго родительского растения аллель «а-малое» содержит мутацию, в результате которой фермент теряет активность и цвет цветков белый.
У растений с генотипом A-большое а-малое только один из аллелей кодирует фермент, который ответственный за пурпурный цвет. Поэтому ферментов вырабатывается мало и цвет цветков получается розовый. По внешнему виду цветка можно определить его генотип.
При неполном доминировании аллельный ген не до конца подавляет действие второго. При этом возникают промежуточные признаки.
Следующий тип взаимодействия неаллельных генов, который мы с вами рассмотрим ― кодоминирование.

При кодоминировании назвать один из аллелей доминантным, а другой — рецессивным нельзя, так как оба аллеля в равной степени влияют на фенотип.
Признаки, за которые отвечает каждый из аллелей, проявляются при кодоминировании одновременно и в полной мере.
Типичный пример кодоминирования — наследование групп крови системы АВ0 (а-бэ-ноль) у человека. При этом А и В-доминантные аллели, а 0-рецессивный. Человек может наследовать эти аллели в следующих комбинациях: 00-первая группа крови, А-большое А-большое и А-большое 0 - вторая, ВВ (читается Бэ-большое) и В-большое 0- третья, А-большое В-большое (когда два доминантных гена определяют признак вместе, не подавляя друг друга) — четвертая группа крови.
То есть при кодоминировании проявляются 2 признака сразу.
Итак, мы рассмотрели типы взаимодействия между аллельными генами, перейдём к взаимодействию между неаллельными генами.
Дополнительное (комплементарное) взаимодействие ― это такой тип взаимодействия неаллельних генов, когда один доминантный ген дополняет действие другого неаллельного доминантного гена и они вместе определяют новый признак, который отсутствует у родителей. Причём соответственный признак развивается только в присутствии обоих неаллельних генов.
Например, при скрещивании двух чистых линий душистого горошка, которые имеют белые цветки, у гибридов первого поколения все цветки будут пурпурными.
Почему так происходит? Дело в том, что в данном случае доминантные неаллельные гены А-большое и Бэ-большое, каждые в отдельности кодируют недостаточное количество пурпурного пигмента для окраски цветка. И только при наличии обоих неаллельных доминантных генов А-большое и Бэ-большое, которые кодируют достаточное количество пигмента цветки окрашиваются в пурпурный цвет.
Следующее взаимодействие неаллельных генов ― эпистаз.
Это вид взаимодействия неаллельных генов, при котором один ген подавляет (не даёт проявиться в фенотипе) другой ген.
Эпистаз противоположен комплементарному взаимодействию, здесь ген не дополняет действие другого, а подавляет.
Гены, которые подавляют действие других генов, называют генами-ингибиторами. Подавление могут вызывать как доминантные, так и рецессивные гены, и в зависимости от этого различают эпистаз доминантный и рецессивный.
Доминантный эпистаз характерен для окраски плодов у тыквы.
У этого растения известно три цвета плодов: белый, жёлтый и зелёный.
Доминантный аллель гена А определяет жёлтую окраску тыквы, рецессивный аллель а-малое — зелёный цвет. Второй ген Бэ-большое проявляет эпистатическое действие — подавляет образование пигмента как жёлтого, так и зелёного цвета, делая их белыми. Рецессивный аллель бэ-малое — не влияет на проявление окраски плодов тыквы.
При скрещивании растений с белыми и зелёными плодами все потомство первого поколения будет белым, так как ген Бэ-большое подавляет действие гена А-большое.
А во втором поколении наблюдается расщепление признаков. В этом случае рецессивная гомозигота (a-малое a-малое бэ-малое бэ-малое) фенотипически отличима от гетерозиготных классов. Таким образом, растения, имеющие в своём генотипе хотя бы один доминантный ген Бэ-большое, будут образовывать белые плоды независимо от того, какие аллели гена А-большое будут присутствовать в их генотипах.
Следующий тип взаимодействия неаллельных генов — полимерное взаимодействие генов.
Полимерия — это вид взаимодействия двух и более неаллельных генов, доминантные аллели которых однозначно влияют на развитие одного и того же признака.
Полимерные гены обозначаются одной буквой, аллели одного локуса имеют одинаковый цифровой индекс.
Действие признаков суммируется, и чем больше в генотипе доминантных пар генов, которые влияют на количественный признак, тем сильнее они проявляются.
Полимерное действие генов может быть кумулятивным и некумулятивным.
При кумулятивной полимерии интенсивность значения признака зависит от суммирующего действия генов: чем больше доминантных аллелей, тем больше степень выраженности признака.
При некумулятивной полимерии количество доминантных аллелей на степень выраженности признака не влияет, а признак проявляется при наличии хотя бы одного из доминантных аллелей.
Приведём пример кумулятивной полимерии.
Бурый цвет зерна пшеницы обусловлен доминантными генами из двух пар аллелей —
А-большое-1 и А-большое-2.
Поэтому самые яркие семена имеют растения, у которых четыре доминантных гена А-большое-1 А-большое-1 А-большое-2 А-большое-2.
У растений с генотипом А-большое-1 а-малое-1 а-малое-2 а-малое-2 или а-малое-1
а-малое-1 А-большое-2 а-малое-2 — зерно окрашено слабо.
А у растений с генотипом а-малое-1 а-малое-1 а-малое-2 а-малое-2 ― зерно не окрашено.

Например, при скрещивании растения с ярко бурыми семенами и растения с неокрашенными семенами в первом поколении все организмы получаются со слабо окрашенными семенами, так как им досталась половина доминантных генов от первого родителя.
При скрещивании гетерозиготных растений из первого поколения происходит вот такое расщепление.
Другой пример полимерного действия генов — наследование окраски кожи человека. Цвет кожи определяется четырьмя генами, расположенными в четырёх различных хромосомах. Эти гены отвечают за синтез тёмного пигмента кожи — меланина.
У европейцев чаще всего встречаются рецессивные гены, отвечающие за синтез меланина. А у самых темных африканцев доминантные.

 1794
1794

